
太阳光提供给植物能量以及波长、周期、强度和方向等信号。高等植物监测日照长度(即光周期)的变化,通过调节开花时间以确保繁殖成功。日照长度由叶片感知后诱导成花素基因FT在维管束的表达,FT编码的成花素由叶片转移至茎顶端分生组织,促进植物开花。长日照下,FT被光周期输出因子CO在韧皮部于黄昏时(dusk)特异激活,CO-FT调控单元是光周期途径的核心调控模式,CO结合在FT靠近转录起始位点的近端启动子区从而激活其表达。
从植物到人类的多细胞真核生物中,PcG因子在关键发育基因的转录抑制中发挥重要作用。PcG复合物本身并不与DNA特异性结合,而是通过Polycomb响应元件(PREs)的募集等方式到达靶基因的染色质。PcG因子抑制基因表达的活性在染色质水平被TrxG因子所拮抗。PcG和TrxG蛋白分别参与基因抑制和激活,通过形成不同的复合物,维持关键发育调控基因的特异性转录。
近年来,随着研究的深入,人们关于光周期开花提出了新的问题,如光周期变化是否造成FT染色质状态的转换,从而塑造FT的表达?中科院植物所林荣呈团队发现叶脉中表达的染色质重塑因子PKL在长日照结束时激活FT。CO和染色质重塑因子PKL间存在遗传和蛋白水平的互作,二者结合FT染色质依赖彼此的功能存在。通过分析转录因子结合FT的调控区域后发现,黄昏时,CO结合FT位点的区域形成染色质“开放”区域,而在晚上该区域相对“关闭”。PKL突变导致该区域在黄昏和晚上都处于相对“关闭”状态 (图1)。因此,PKL对于光周期调控FT染色质激活和抑制状态的转变是必需的。
PcG蛋白介导的对FT的表观抑制可阻止植物提早开花,在FT的表达调控中发挥重要作用。目前,人们对FT表观抑制的触发机制并不清楚。林荣呈团队在FT的第二个内含子到3'UTR之间鉴定到H3K27me3组蛋白修饰高水平富集区域(HTR),该区域存在两个紧密相邻的RY元件,该元件介导了对转基因植物的开花抑制,能够被转录抑制因子VAL1所识别。同时,VAL1蛋白募集PcG蛋白LHP1和MSI1至FT染色质,在HTR处建立H3K27me3,在黄昏前和夜晚直接抑制FT表达(图1)。因此,DNA顺式元件通过招募VAL1介导FT的表观抑制。
PcG和TrxG蛋白拮抗调控关键基因以维持发育的时空特性在动物中被广泛报道,在植物中却研究较少。TrxG因子能否直接激活FT表达?TrxG和PcG因子能否拮抗调控FT的表达?其详细机制仍有待阐明。林荣呈团队研究发现:由于FT异位高表达,PcG的拟南芥突变体有明显的早花表型,将编码TrxG(类)因子如染色质重塑因子PKL 和COMPASS蛋白ATX1的基因突变后导入其中, FT的表达几乎回复至野生型水平和模式。PKL与ATX1能够相互作用,介导FT位点H3K4的三甲基化,从而拮抗PcG蛋白对FT表达的抑制作用(图1)。该研究揭示了TrxG(类)蛋白拮抗PcG介导的对FT的转录抑制,该机制维持了FT的特异且适当表达,从而调控光周期开花。
上述研究成果揭示了成花素基因FT表观遗传调控的分子机制,分别于2019年8月在Plant, Cell & Environment正式发表,于2019年7月和8月在Plant Physiology在线发表。林荣呈研究组的景艳军副研究员为第一作者,林荣呈和景艳军为通讯作者。这些研究得到了国家重点研发计划、国家自然科学基金和中科院青促会专项等项目的资助。
文章链接:
https://onlinelibrary.wiley.com/doi/full/10.1111/pce.13557
http://www.plantphysiol.org/content/early/2019/07/09/pp.19.00642.long
http://www.plantphysiol.org/content/early/2019/08/03/pp.19.00596.long
(光合实验室供稿)
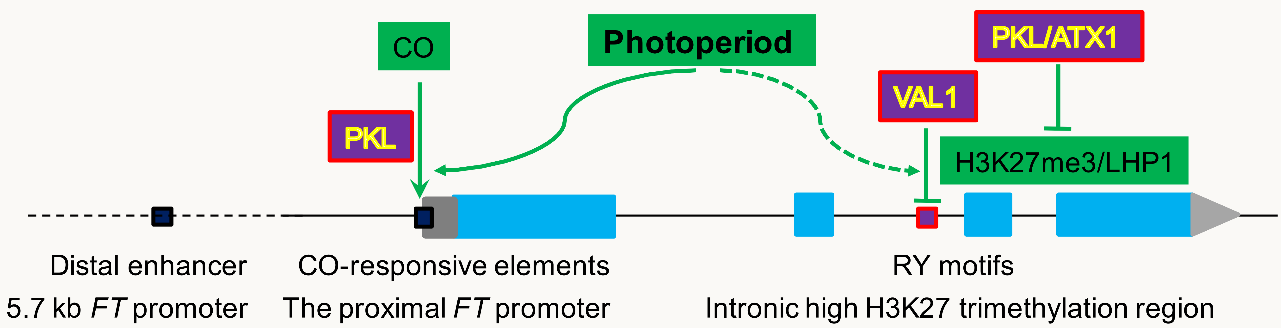
图1、成花素基因FT表观遗传调控的分子机制
红色框/紫色背景标注的蛋白因子或DNA元件是本研究关注的内容

