
2005年版《中国药典》将忍冬科忍冬属植物忍冬(Lonicera japonica Thunb.)规定为中药材金银花的唯一植物来源。然而,我国历史及传统上作为中药金银花流通及使用的忍冬属植物却不限于此。其中,灰毡毛忍冬(L. macranthoides Hand.-Mazz.)在中医药临床应用与中医制剂中一直作为金银花使用。尽管灰毡毛忍冬也被编入《中国药典》,但其被归入“山银花”,而各种中药配方一直沿用“金银花”,导致我国南方数省出产的灰毡毛忍冬很快就失去了中药材市场,给农民造成了巨大的经济损失。灰毡毛忍冬能否继续作为中药材金银花来使用?争论的焦点在于忍冬和灰毡毛忍冬中的药效成分之一“木犀草苷”含量的高低及其成因。中科院植物研究所王亮生研究组以忍冬和灰毡毛忍冬‘湘中三号’为研究材料,就这一问题进行了深入探讨。
研究人员发现,负责木犀草素合成的关键基因——细胞色素P450家族的黄酮合酶基因(FNS)在忍冬和灰毡毛忍冬中存在较大差异。从忍冬中克隆得到的2个黄酮合酶基因(LjFNSII-1和LjFNSII-2)各编码2个定位在内质网上的蛋白(LjFNSII-1.1&1.2、LjFNSII-2.1&2.2)。其中,仅LjFNSII-1.1和LjFNSII-2.1具有催化活性,且这两个蛋白仅有1个氨基酸的差异。同源建模以及定点突变酶活试验证明,在242位的碱性氨基酸更有利于维持蛋白α-helix的稳定性,从而使LjFNSII-1.1具有较高的催化活性。灰毡毛忍冬中的FNS仅编码一个有活性的蛋白LmFNSII-1.1。酵母异源表达的微粒体蛋白酶活实验表明,该蛋白的催化活性远低于忍冬中的黄酮合酶。蛋白与底物之间的分子对接模拟实验以及定点突变实验表明,位于381位的亮氨酸和206位的甲硫氨酸对黄酮合酶的活性起着重要作用。上述3个基因(LjFNSII-1.1、LjFNSII-2.1和LmFNSII-1.1)的表达模式分析和转基因验证实验的结果最终揭示,灰毡毛忍冬中的黄酮合酶基因LmFNSII-1.1在花中的低表达量及其较低的酶活性,是导致其花蕾中木犀草苷含量低的原因。
以灰毡毛忍冬为种源形成的一系列品种,其花蕾数量众多、花蕾聚合成伞状或团状花序、花冠整齐、发育时期一致且花期集中、花朵几乎不开放、可一次性采摘,在生产上可大大节约采摘时间和劳动成本,产量显著高于忍冬由来的品种。今后,通过分子育种手段定向提高灰毡毛忍冬花中的木犀草苷含量,将有望作为忍冬(金银花)的优良替代品。该研究成果对灰毡毛忍冬及其品种的品质改良和新品种创制具有深远意义。
该研究成果于2016年1月12日在线发表在Scientific Reports期刊上。研究组博士生吴杰为论文的第一作者。研究得到了科技部基础性工作专项“武陵山区生物多样性综合科学考察”的资助。
论文链接:
http://www.nature.com/articles/srep19245
(资源植物实验室供稿)
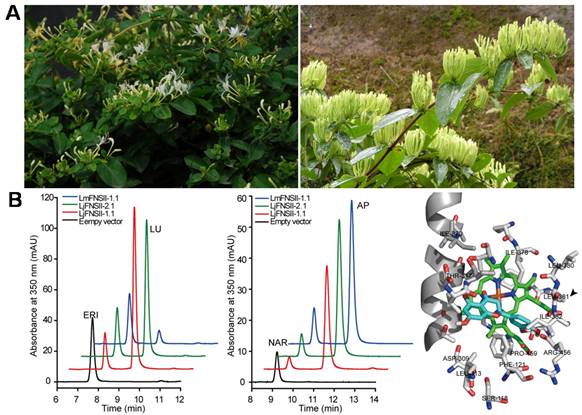
(A) 图左为忍冬,图右为灰毡毛忍冬‘湘中三号’;(B) 图左、图中分别为酵母细胞表达LjFNSII-1.1&2.1的提取物的HPLC图谱(350 nm),图左显示eriodictyol(ERI)直接转化为luteolin(LU),图中显示naringenin(NAR)直接转化为apigenin(AP);图右为LjFNSII-1.1的酶活性中心。

